本帖最后由 荷花池荒岛 于 2014-5-4 01:07 编辑
http://www.intechopen.com/books/ ... lignant-mesotheliom
2.2. EPIDERMAL GROWTH FACTOR RECEPTOR
The epidermal growth factor receptor (EGFR) is the cell-surface receptor for members of the epidermal growth factor family (EGF-family) of extracellular protein ligands (Herbst, 2004). Upon activation by its growth factor ligands, EGFR undergoes a transition from an inactive monomeric form to an active homodimer. In addition, EGFR may pair with another member of the ErbB receptor family, such as ErbB2/Her2/neu, to create an activated heterodimer. EGFR dimerization stimulates its intrinsic intracellular protein-tyrosine kinase activity. As a result, autophosphorylation of several tyrosine residues in the C-terminal domain of EGFR occurs (P-EGFR). This autophosphorylation leads to the activation of downstream signalling cascades including the RAS/extracellular signal regulated kinase (ERK) pathway, the phosphatidylinositol 3-kinase/AKT (PI3K/AKT) pathway and the Janus kinase/Signal transducer and activator of transcription (JAK/ STAT) pathway (Fig.3).

These pathways act in a coordinated manner to promote cell survival (Oda et al., 2005). Such proteins modulate phenotypes such as cell migration, adhesion, and proliferation. EGFR is reportedly over-expressed in a wide variety of malignancies. Various studies suggest that receptor tyrosine kinase activation participates in the oncogenic progression of non neoplastic mesothelial progenitor cells to malignant mesothelioma. Asbestos fiber interact with the external domain of the EGFR to cause dimerization, activation and increased EGFR mRNA and protein levels in rat and human SV-40 immortalized mesothelial cells (Shukla et al., 2011). Up-regulated EGFR and resulting tyrosine phosphorylation leads to the Ras activation which phosphorylates directly and activates Raf (Rapidly Accelerated Fibrosarcoma). Raf is responsible for phosphorylation of the mitogen associated / extracellular regulated kinase-1 (MEK) which in turn phosphorylates extracellular regulated kinases (ERK) on specific residues of threonine and tyrosine (Ras-Raf-MEK-ERK mitogen activated protein kinase (MAPK) pathway). ERK activates a variety of substrates involved in cell cycle. The ERK family consists of at least seven isoforms, and little is known about their regulation and function. ERK1/2 phosphorylation by asbestos, is dependent on phosphorylation of the EGFR. Moreover, has been shown that ERK5, a redox-sensitive kinase known to mediate c-jun proto-oncogene expression is activated by asbestos. ERK1/2 and ERK5 are all important in asbestos-induced proliferation and this may be the result of increases in the mRNA levels of AP-1 family members. The ERK5 pathway may be contributing selectively to the regulation of c-jun, whereas ERK1/2 pathways may regulate c-fos, fra-1 and c-jun. Has been linked ERK1/2-dependent fra-1 expression to mesothelial cell transformation by asbestos and the protracted expression of this gene may be a result of initial increases in c-fos and c-jun (Scapoli et al., 2004). The phosphoinositide 3-kinase (PI3K)/AKT pathway, plays a critical role for the cell cycle progression in human MM cells [ Altomare et al.,2005). AKT, and the downstream mTOR are involved in cell growth and survival, and they are often found to be activated in MM (Carbone et al., 2012). It was reported previously that STAT1 and STAT3 are deregulated MM (Kothmaier et al., 2008).The JAK/STAT signalling pathway is the principal signalling mechanism for growth factors in mammals. JAK activation induces a variety of biological responses such as cell proliferation, diff erentiation and cell migration. In addition, MM cell lines are reported to express EGFR and transforming growth factor-α (TGF-α), suggesting an autocrine role for EGFR in MM (Cai et al., 2004; Jänne et al., 2002). EGFR immunopositivity has been indicated as a poor prognostic factor in many solid tumors in the past (Nicholson et al., 2001). The EGFR expression in MM has been previously reported, with controversial results, possibly due to the lack of standardized method for EGFR detection and quantification (Dazzi et al., Destro et al., 2006; 1990; Govindan et al., 2005; Ramael et al., 1991; Trupiano et al., 2004). Until now, the role of immunohistochemistry (IHC) EGFR positive staining in influencing prognosis of MM is not clear. Some authors did not find differences in survival when IHC EGFR positive or negative staining were compared (Destro et al., 2006; Okuda et al., 2008). This is because only few reports analyzed the effect of IHC EGFR positive status and cell subtype in MM patients. Recently EGFR overexpression is identified by IHC in 52% of epithelial MM and is demonstrated to be a factor negatively affecting prognosis (Rena et al., 2011). In view of these studies, EGFR was targeted for MM therapy, but despite the high expression of EGFR not all cells are sensitive to EGFR inhibitors (Garland et al., 2007). Many efforts are now directed to understand the lack of sensitivity of MM to EGFR inhibitors. In one such study, EGFR mutations were found in 31% (9 of 29) of malignant mesothelioma cases. Seven of these mutations were novel, and one was the L858R mutation described in NSCLC (Foster et al., 2009). Activating EGFR mutations in MM associated with optimal resectability and prolonged survival. Clinically these mutations may ultimately have utility in patient selection for surgery, systemic therapy, and selection for EGFR-TKI (tyrosine kinase inhibitor). The clinical course of MM patients with EGFR mutant tumors appear to share same 'relative' improved clinical outcome like mutant EGFR-NSCLC (Foster et al., 2010. Study shows the ineffectiveness of the EGFR inhibitors due to coactivation of multiple receptor tyrosine kinase (EGFR, ERBB3, MET, and AXL) in individual mesothelioma cell lines (Ou et al., 2011) Thus, a combination therapy, could be a winning strategy in the treatment of mesothelioma.
|
 各位老师,帮忙看看我的问题,求帮助
父亲去年2023年2月份体检发现肺癌 晚期,免疫治疗一年,用药欧迪沃 期间病灶一直稳定
各位老师,帮忙看看我的问题,求帮助
父亲去年2023年2月份体检发现肺癌 晚期,免疫治疗一年,用药欧迪沃 期间病灶一直稳定
 右额上回内缘、左中央前回前缘皮质表
这是最近两次磁共振的结果,请大家帮忙看看:右额上回内缘、左中央前回前缘皮质表面微
右额上回内缘、左中央前回前缘皮质表
这是最近两次磁共振的结果,请大家帮忙看看:右额上回内缘、左中央前回前缘皮质表面微
 EXTENTORCH研究:最亲民的小细胞肺癌
小细胞肺癌约占肺癌的15%左右[1-2],是肺癌中恶性程度高、侵袭性强的一类亚型。与发病
EXTENTORCH研究:最亲民的小细胞肺癌
小细胞肺癌约占肺癌的15%左右[1-2],是肺癌中恶性程度高、侵袭性强的一类亚型。与发病
 新人,肺腺癌晚期 多发骨转移 有病理
母亲今年5月份确诊肺腺癌晚期,原先体感还好就是后背痛,不能侧翻,不能好好睡觉,目
新人,肺腺癌晚期 多发骨转移 有病理
母亲今年5月份确诊肺腺癌晚期,原先体感还好就是后背痛,不能侧翻,不能好好睡觉,目
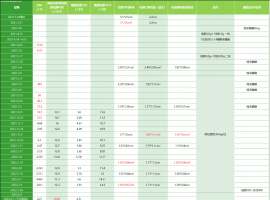 阿法替尼13个月,达克替尼5个月,目
2021年3月的最后一天,很平常的一天,一份CT报告打破了这平静的生活!也许再过一天,
阿法替尼13个月,达克替尼5个月,目
2021年3月的最后一天,很平常的一天,一份CT报告打破了这平静的生活!也许再过一天,





 显身卡
显身卡