| |
|
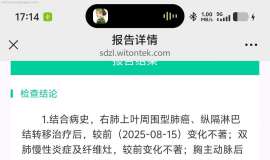
一切恩爱会,无常难得久;
生世多畏惧,命危于晨露。 由爱故生忧,由爱故生怖; 若离于爱者,无忧亦无怖。 |
|
| |
|
一切恩爱会,无常难得久;
生世多畏惧,命危于晨露。 由爱故生忧,由爱故生怖; 若离于爱者,无忧亦无怖。 |
|
| |
|
一切恩爱会,无常难得久;
生世多畏惧,命危于晨露。 由爱故生忧,由爱故生怖; 若离于爱者,无忧亦无怖。 |
|
| |
|
一切恩爱会,无常难得久;
生世多畏惧,命危于晨露。 由爱故生忧,由爱故生怖; 若离于爱者,无忧亦无怖。 |
|
| |
|
一切恩爱会,无常难得久;
生世多畏惧,命危于晨露。 由爱故生忧,由爱故生怖; 若离于爱者,无忧亦无怖。 |
|
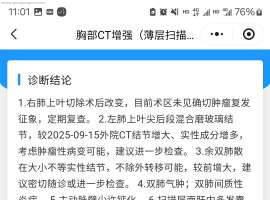 肺腺癌术后不到两个月快速进展,求教
25年9月底,手术切除了右上肺,有两个淋巴转移,分期是2期。后续在进行辅助治疗的时候
肺腺癌术后不到两个月快速进展,求教
25年9月底,手术切除了右上肺,有两个淋巴转移,分期是2期。后续在进行辅助治疗的时候
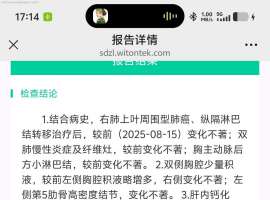 不知不觉进入第七年。。胸腔积液怎么
中间这几年很顺利也很稳定 这几年都没再关注这个平台。空窗一年半,最近一次复查胸腔
不知不觉进入第七年。。胸腔积液怎么
中间这几年很顺利也很稳定 这几年都没再关注这个平台。空窗一年半,最近一次复查胸腔
 75岁老人肺癌有转移情况,是选择积极
还没有做 穿刺和基因检测,家里做医生的亲戚建议我姑息治疗,怕我人财2空
毕竟有75岁
75岁老人肺癌有转移情况,是选择积极
还没有做 穿刺和基因检测,家里做医生的亲戚建议我姑息治疗,怕我人财2空
毕竟有75岁
 流感高峰期,肿瘤患者要注意!抗病毒
作者:seacat
“我以为就是感冒,结果三天后高烧不退,还喘不上气。”
这是王阿姨(化
流感高峰期,肿瘤患者要注意!抗病毒
作者:seacat
“我以为就是感冒,结果三天后高烧不退,还喘不上气。”
这是王阿姨(化
 一位肿瘤科护士的硬核抗癌路:4次基
讲述者:阿雷一点都不累整理者:Tony
2023年12月28日那一天,我34岁的妹妹被确诊为Ⅳ
一位肿瘤科护士的硬核抗癌路:4次基
讲述者:阿雷一点都不累整理者:Tony
2023年12月28日那一天,我34岁的妹妹被确诊为Ⅳ





 寄予极大期望的易要不要再坚持吃段时间?才7天,可是感觉每天都在加重病情。明明,免疫组化是中阳啊!为什么要去测该死的突变,导致犹豫着始终不敢用。。是扩散太多了吗?失控了吗?感觉左眼周似乎有小的丘疹出现?加量吃么?还是换299804+184?
寄予极大期望的易要不要再坚持吃段时间?才7天,可是感觉每天都在加重病情。明明,免疫组化是中阳啊!为什么要去测该死的突变,导致犹豫着始终不敢用。。是扩散太多了吗?失控了吗?感觉左眼周似乎有小的丘疹出现?加量吃么?还是换299804+184? 显身卡
显身卡 / |1 C- U5 q$ W1 D3 l2 U% O
/ |1 C- U5 q$ W1 D3 l2 U% O